
1986年在北京大学生物系获学士学位。1991年在北京大学生物系获博士学位。1991-1994年,中科院遗传所工作。1994-1997年,美国堪萨斯州立大学博士后研究。1998-2005年,中科院遗传发育所副研究员xhjc1新黄金城。2006年,中科院遗传发育所研究员。2009年获得国家杰出青年基金资助。主持或参与多项国家自然科学基金项目,转基因专项,973及先导专项等。获得基因发明专利30余项,发表SCI论文100余篇。
有关乙烯的相关背景和知识请参考曹务强博士在科学网撰写的系列博文,他会带你进入一个精彩而奇妙的植物世界。
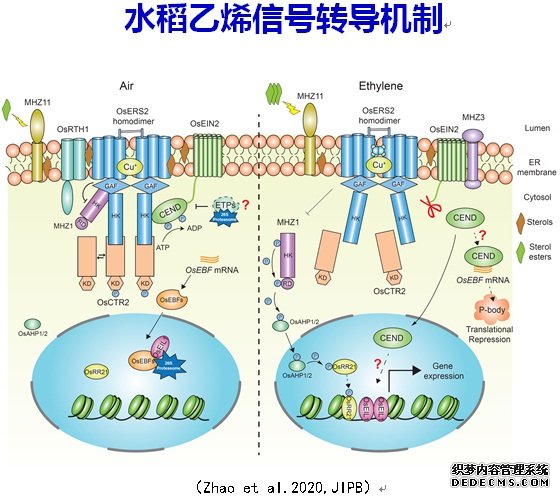
乙烯对于水稻的适应性和生长发育具有重要作用。利用水稻乙烯‘二重’反应,即水稻黄化苗的根生长受乙烯抑制而地上部胚芽鞘的生长受到促进,筛选到了一系列水稻乙烯反应突变体编码一个组氨酸激酶OsHK1,MHZ1/OsHK1可进行自体磷酸化,并通过磷酸传递蛋白OsAHP1和OsAHP2将磷酸基团传递给下游的响应因子OsRR21。乙烯受体MHZ12/OsERS2和其它乙烯受体可以通过其GAF结构域与MHZ1互作抑制MHZ1的激酶活性,进而抑制MHZ1介导的磷酸传递系统。在乙烯存在下,乙烯通过抑制受体功能从而解除了受体对MHZ1的抑制,激活MHZ1介导的磷酸传递和根乙烯反应。E3泛素连接酶a可以与非典型Aux/IAA蛋白OsIAA26结合,通过泛素化降解OsIAA26蛋白,调控乙烯对水稻根伸长的抑制作用。内质网定位的MHZ3通过与OsEIN2蛋白的N端跨膜区Nramp-like结构域结合,抑制OsEIN2的泛素化,稳定OsEIN2蛋白,从而保护OsEIN2蛋白不被蛋白酶体途径降解维持乙烯反应。这项研究强化了OsEIN2的N端跨膜区在乙烯信号转导中的作用。编码一个GDSL类型的脂酶,具有脂酰水解酶活性。乙烯处理后,MHZ11可能通过水解磷脂而产生脂肪酸,脂肪酸与甾醇形成甾醇酯,降低了膜上微域(microdomain)的甾醇含量,从而减弱了微域中受体和OsCTR2互作及后者磷酸化,进而启动了下游乙烯信号转导。我们的研究揭示了MHZ11通过影响甾醇代谢平衡及受体-OsCTR2互作和OsCTR2磷酸化来激活下游乙烯信号转导的新机制。前期我们研究发现MHZ7/OsEIN2和MHZ6/OsEIL1与拟南芥的乙烯信号转导组分EIN2和EIN3类似,也是水稻乙烯信号转导的关键组分。转录因子MHZ6/OsEIL1和其同源蛋白OsEIL2分别调控水稻黄化苗根和胚芽鞘的乙烯反应;水稻乙烯信号转导途径通过促进Na吸收导致盐敏感反应;乙烯途径通过MHZ5/类胡萝卜素异构酶介导的类胡萝卜素合成途径和MHZ4/OsABA4介导的ABA合成途径调控根的生长。这些机制与拟南芥中的完全不同。在拟南芥中,转录因子EIN3和EIL1对乙烯反应的调控没有器官特异性;乙烯信号转导提高耐盐性;ABA通过乙烯途径调控根的生长。对一个)突变体的分析发现,乙烯途径抑制GY1/磷脂酶A1介导的茉莉酸合成促进中胚轴和胚芽鞘伸长;也鉴定了一个优异等位变异
大豆是重要的蛋白和油料作物,我们致力于鉴定相应的调节基因并研究其在调控大豆油脂等品质和胁迫反应中的功能。这些研究对于改善大豆品质及耐逆性具有重要的实际意义。通过对10个栽培大豆资源和10个野生大豆资源不同发育阶段籽粒进行RNA测序,鉴定了2个栽培大豆特异的基因共表达调控网络。通过比较网络节点基因与相应籽粒性状QTL的重叠性,鉴定出了调控大豆油脂合成的基因,其中GmZF351是调控从糖酵解、脂肪酸合成、TAG形成,到油体形成的重要因子(master regulator)。进一步利用源于高油低油材料的RIL群体,通过重测序及QTL定位,鉴定了油脂和粒重调控QTL,并克隆了来源于野生大豆的基因,该基因编码的蛋白通过与GmBZR1互作使其脱磷酸化,从而激活BR途径提高粒重。在栽培大豆中还有约40%材料不含PP2C-1位点,进一步将此位点导入将可能提高大豆产量潜力。前期通过对大豆不同器官及发育中的大豆籽粒进行转录组分析,鉴定了数十个种子特异表达且与油份积累同步的转录因子基因。深入研究发现大豆GmbZIP123可通过激活蔗糖转运蛋白基因和细胞壁蔗糖转化酶基因表达,从而促进糖分从叶片向种子的转运来提高油脂合成和积累。也发现了另一个大豆GmMYB73可与GL3和EGL3互作抑制的抑制。PLDα1高表达促进磷脂酰胆碱PC转化为磷脂酸PA和二脂酰基甘油DAG和三酰甘油TAG,使种子和叶片油份积累得到提高。我们的研究揭示了大豆油份积累的新机制,对于大豆品质改良具有重要意义。也鉴定了一系列胁迫应答的转录因子等调控基因,研究了它们在大豆耐逆中的作用。